|
Гипермаркет знаний>>Естествознание>>Естествознание 10 класс>> Кайнозой: наступление криоэры. Новые типы сообществ – тропические леса и травяные биомы. Эволюция млекопитающих и появление человека
Кайнозой: наступление криоэры. Новые типы сообществ – тропические леса и травяные биомы.
Эволюция млекопитающих и появление человека.
Мезозой, как мы помним из главы 9, был термоэрой и отличался «райским» климатом: теплым и выровненным по всей Земле, с ослабленным температурным градиентом между тропиками и полюсом. В рамках термоэр неоднократно происходили свои похолодания (например, в маастрихте), однако масштабы даже крупнейших из них не идут ни в какое сравнение с ледниковыми эпохами криоэр. В самом начале кайнозоя (в палеоцене и эоцене) климатическая ситуация оставалась еще прежней, мезозойской, а затем, в олигоцене, начались изменения, приведшие в конце концов к... так и тянет сказать: «к тому самому климатическому безобразию, в каковом мы теперь имеем счастье прозябать (в буквальном смысле этого слова)». Одной из главных причин этих изменений считают перестройку глобальной системы морских течений, вызванную дрейфом континентов, что затруднило теплообмен между полярными и экваториальными бассейнами.
Согласно моделям климатологов, наиболее интенсивный теплообмен будет достигнут при таком взаиморасположении материков и полюсов, когда планета имеет свободные от суши полюса и экватор (вдоль которого складывалась бы под действием кориолисовых сил единая экваториальная циркуляция). Меридионально развернутые материки, расположенные в средних широтах, отводят рукава экваториальной циркуляции в высокие широты, откуда вдоль противоположных их берегов возвращается в экваториальную зону холодная вода из полярных бассейнов – возвращается сразу, не застаиваясь в полярных циркуляциях. Как легко видеть (рисунок 53, а), мезозойская ситуация была в этом смысле близка к идеальной.
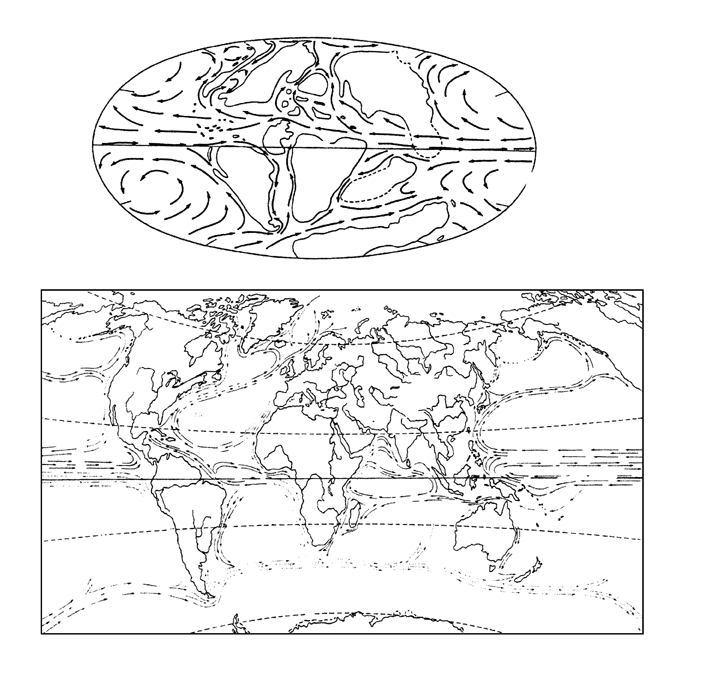
РИСУНОК 53. Материки и морские течения. Мезозойское (средний мел) (а) и современное (б) расположение материков.
В кайнозое картина становится близка к обратной, и причиною тому – перемещения материков, на которые раскололся суперконтинент Гондвана (рисунок 53, б). К концу эоцена движущаяся на север Индия натыкается на Азию, а кора в месте их столкновения сминается в Гималаи. Чуть позже в Евразию втыкается с юго запада Африка (с еще не отделившейся от нее Аравией) – в результате поднимаются горные хребты южной Европы и Иранское нагорье. Океан Тетис, отделявший в мезозое северные (лавразийские) материки от южных (гондванских) – а именно по нему тогда шла экваториальная циркуляция – исчезает; ныне от Тетиса остались лишь изолированные морские бассейны (Средиземное, Черное и южная часть Каспийского моря), не имеющие постоянной связи с мировым океаном . В южном полушарии Антарктида последовательно отрывается от Австралии (эоцен) и Южной Америки (олигоцен), «наползает» на Южный полюс, а вокруг нее складывается замкнутая антарктическая циркуляция; все это приводит к тому, что в олигоцене в Трансантарктических горах возникают первые ледники, которые в миоцене покрывают единым ледяным щитом весь материк. В плиоцене возникает Панамский перешеек – соединяющий Южную Америку с Северной, но разделяющий Атлантический и Тихий океаны, – и глобальная экваториальная циркуляция оказывается полностью разрушенной (ныне мы имеем лишь один ее сегмент – в центральной части Тихого океана). Разрушение экваториальной циркуляции, возникновение «антарктического холодильника» на Южном полюсе и замыкание северного полярного бассейна – все это приводит тому, что в четвертичном периоде (в плейстоцене) на материках Северного полушария развиваются грандиозные покровные оледенения (см. главу 14).
Надо заметить, что отсутствие в мезозое холодных климатических зон давно не вызывает сомнения у палеоклиматологов. Однако они зачастую не обращают внимания на то, что при более равномерном, чем ныне, распределении тепла по поверхности планеты не должно было существовать и климата типа современного тропического. Но тогда и сообщества, сложившиеся в условиях не существовавших ранее типов климата (бореального с одной стороны и тропического – с другой), должны быть эволюционно наиболее молоды... Если в некий момент на полюсах стало «слишком холодно», а на экваторе – «слишком жарко», то реликты предыдущей эпохи имеют наилучшие шансы выжить прежде всего в субтропиках, господствовавших ранее по всей Земле. Специальный анализ, проделанный В.В. Жерихиным (1978), показал, что так оно и есть. Судя по всему, некоторые субтропические сообщества (такие, как, например, леса из южного бука в Новой Зеландии, южной Австралии и андийской Южной Америке) сохранили не просто отдельные реликты раннего кайнофита, а самую структуру раннекайнофитных биоценозов.
Что же касается тропических лесов (мы будем называть их южноамериканским словом гилея), то укоренившиеся представления о глубокой древности их биоты являются очередным «научным предрассудком»; они связаны в основном с тем, что понятие «тропики» зачастую употребляют нечетко. В астрономических тропиках – т.е. между тропиками Рака и Козерога – действительно есть сообщества с весьма архаичными элементами (например, индонезийские «туманные леса» из древовидных папоротников), однако все они приурочены к горам, и существуют, на самом деле, в условиях субтропического или даже умеренного климата. Если же рассмотреть лишь климатические тропики, т.е. экосистемы, реально существующие в условиях тропического климата (с температурой самого холодного месяца не менее 18°С) – гилею и саванны, то окажется, что их биота практически лишена архаичных черт. Мы уже упоминали (глава 12) об отсутствии в составе тропических лесов голосеменных, столь характерных для мезозойских сообществ; ныне те приурочены либо к субтропикам (саговники, араукарии, таксодиум), либо к умеренной зоне (секвойя, гинкго).
Подсчеты Жерихина показали, что практически все по настоящему древние группы насекомых (такие, как скорпионницы или сетчатокрылые) избегают тропиков. Общее число семейств, не встречающихся в истинных, климатических, тропиках очень велико, тогда как чисто тропических (в этом смысле) семейств среди насекомых – как это ни удивительно – ничтожно мало. Разумеется, тропическая энтомофауна чрезвычайно богата, и есть множество преимущественно тропических семейств – однако почти все они эволюционно молоды и не обнаружены в ископаемых донеогеновых фаунах, либо представлены в них очень скудно.
Еще более интересны в этом плане приводимые Жерихиным (1980) данные по экологической структуре тропических лесов:
"Сообщества типа современной гилеи с их сложной ярусной структурой и высокой степенью полидоминантности вообще не могут существовать без участия филогенетически молодых групп. Полидоминантность в вечнозеленых лесах [в отличие от листопадных – К.Е.] поддерживается только наличием специализированных опылителей (пчелы, антофильные птицы и рукокрылые), поскольку анемофилия эффективна лишь для самых высокоствольных деревьев...
Деструкция органического вещества как в гилейных, так и в саванновых сообществах производится прежде всего термитами... Препятствуя образованию подстилки и выраженного гумусового горизонта (и тем самым – развитого травяного яруса) они, по видимому, обусловили превращение многих травянистых растений в эпифиты и формирование эпифитных консорций, очень типичных для гилеи. Распределяя органическое вещество в мощном слое почвенного профиля более или менее равномерно, они обеспечивают возможность развития корневых систем на самых различных уровнях, в том числе и глубоких, что позволяет существовать гигантским деревьям с глубоко погруженными корнями...
В вечнозеленых лесах, где листья, не сменяясь, существуют на дереве по нескольку лет, требуется эффективное предотвращение сильного повреждения листвы фитофагами; в противном случае деятельность листового насоса не сможет обеспечить существование дерева. Функцию подавления численности открытоживущих фитофагов выполняют чрезвычайно обильные в гилее муравьи..." Все перечисленные выше группы, «определяющие лицо» тропических экоситем – и опылители, и термиты, и муравьи – эволюционно очень молоды и не характерны для раннекайнофитных сообществ. Все это и привело Жерихина к заключению, что тропические сообщества – наравне с бореальными – принадлежат к числу самых молодых на Земле; судя по всему, они возникли не раньше эоцена, а окончательное их формирование произошло уже в неогене.
С уверенностью говорить о времени формирования гилейных экосистем трудно из за очень слабой палеонтологической изученности современной тропческой зоны. Что же касается бореальных сообществ – например, криофильных (холодолюбивых) лесов с доминированием хвойных, типа тайги, – то они, по всей видимости, сложились в палеогене как высотный пояс в горах, а при неогеновом похолодании широко распространились на равнинах Северного полушария. В палеонтологической летописи они впервые достоверно появляются в верхнем миоцене Канадского архипелага (остров Миен); их состав напоминал современные лесотундровые редколесья.
Еще одна характерная для кайнозоя черта – травяные биомы (типа степей и саванн), существование которых основано на коэволюции злаков и крупных травоядных млекопитающих: при ряде условий (в засушливых и малоплодородных районах) животные способны воспрепятствовать восстановлению лесной растительности. В доэоценовые времена, судя по пыльцевым спектрам, сообществ такого типа на Земле не было. По мнению Жерихина (1993), эти биомы первоначально представляли собою сериальные сообщества, ведущие к соответствующим древесным климаксам (стадии с господством трав – например, луга – имеются во всех современных сукцессионных системах), однако затем возникла уникальная ситуация: фитофаги, изымая прирост биомассы (в современных степях это изъятие доходит до 60% растительной продукции – абсолютный рекорд), оказались способны остановить экогенетическую сукцессию на безлесой стадии. Дальнейшая коэволюция млекопитающих и трав привела к стабилизации этих своеобразных «зоогенных климаксов» : необходимый для климаксного сообщества нулевой баланс по органике обеспечивается в них именно деятельностью животных фитофагов. Основные черты, отличающие эти травяные климаксы и от сериальных травяных сообществ, и от лесов – исключительно высокая продуктивность, очень быстрый оборот органического вещества и наличие легко мобилизуемого запаса этой органики в необычайно плодородных почвах (типа черноземов).
Известно, что основой существования этих сообществ является замечательная коадаптация доминирующих в растительном покрове злаков и фитофагов: злаки не имеют эффективной защиты от поедания (вроде колючек, ядовитости и т.п.), зато способны компенсировать обгрызание надземных частей резким ускорением их регенерации. Поэтому регуляция в таком сообществе достигается предельно просто: при снижении пресса фитофагов первичная продукция тоже автоматически снижается – и наоборот. Основную роль в пастбищных цепях этих экосистем играют млекопитающие, прежде всего – различные копытные (потомки кондиляртр – рисунок 55, а) и хищные (потомки креодонтов – рисунок 56, а); об эволюции сообщества позвоночных – чуть далее. Иное дело – детритные цепи. Возникновение стабильных травяных сообществ было бы просто невозможно без появления целого комплекса насекомых копрофагов, перерабатывающих огромную массу экскрементов травоядных . В более древних (палеогеновых) лесных сообществах, где млекопитающие не достигали такой биомассы, как пастбищные копытные, с этой работой справлялись низшие навозники. Важнейшие же современные группы копрофагов (высшие навозники и навозные мухи) в заметных количествах появляются в захоронениях с конца олигоцена. В это же время складывается и не менее важный комплекс насекомых некрофагов (мясные мухи и жуки мертвоеды), ответственный за эффективную переработку трупов травоядных млекопитающих.
Сообщество наземных позвоночных в кайнозое развивалось независимо на трех разобщенных территориях, фаунистические контакты между которыми практически отсутствовали. Австралия (с ее сумчатыми и однопроходными) изолирована и поныне, а Южная Америка сохраняла свою обособленность от остальной суши вплоть до плиоцена, когда возник Панамский перешеек; отсюда и проистекает современное разделение мира на три зоогеографические области: Нотогея (Австралия), Неогея (Южная Америка) и Арктогея (Евразия, Африка и Северная Америка). Так вот, по мнению Жерихина (1993), во всех этих трех областях травяные биомы возникали независимо, на базе совершенно различных комплексов крупных млекопитающих; собственно говоря, есть серьезные основания полагать, что млекопитающие по настоящему вышли в крупный размерный класс лишь в травяных биомах.
Раньше всего (в среднем эоцене) этот процесс начался в Южной Америке. Там среди исходно листоядных «южноамериканских копытных» возникают первые травоядные формы, а также появляются гигантские травоядные броненосцы глиптодонты, смахивающие на небольшой танк (рисунок 54, а). В среднем же эоцене в Южной Америке впервые обнаруживаются и пыльцевые спектры с высоким содержанием пыльцы злаков, палеопочвы степного типа, а также фоссилизированные навозные шары, принадлежащие жукам навозникам. Позднее, в олигоцене и особенно в миоцене, здесь возникает в высшей степени своеобразный комплекс пастбищных травоядных. Он включал неполнозубых (глиптодонтов и наземных ленивцев), «южноамериканских копытных» (различные литоптерны демонстрирует сильное конвергентное сходство частью с лошадьми, частью с верблюдами, у пиротериев много общего со слонами, а среди нотоунгулят были формы, схожие и с носорогами, и с бегемотами, и с кроликами – риснок 54, б г), а также гиганских кавиморфных грызунов (некоторые из этих родственников морской свинки достигали размеров носорога) и просуществовал вплоть до установления в плиоцене сухопутной связи с Северной Америкой.

РИСУНОК 54. Третичные млекопитающие Южной Америки: неполнозубые (а), «южноамериканские копытные» (б г) и хищные сумчатые (д е). (а) – глиптодонт, (б) – литоптерна Thoatherium, (в) – литоптерна Toxodon, (г) – нотоунгулят Macrauchenia, (д) – «сумчатый волк» Prothylacynus, (е) – «сумчатый саблезубый тигр» Thylacosmilus; черепа Thylacosmilus (ж) и «северной» саблезубой кошки Smilodon (з) (в одном масштабе).
Что же касается хищников, то они в древней южноамериканской фауне всегда были в дефиците. Ни один из здешних отрядов плацентарных по не вполне понятным причинам так и не дал плотоядных форм – эту роль исполняли исключительно сумчатые. Довольно разнообразные боргиениды несколько напоминали собак (а еще больше – тилацина, тасманийского «сумчатого волка»), а тилакосмилюс вполне заслуживает названия «сумчатый саблезубый тигр» и являет собою поразительный пример конвергенции с саблезубыми кошками Северного полушария (рисунок 54, д е). Дефицит маммальных хищников (на «несбалансированность» здешних фаун обратили внимание А.С. Раутиан и Н.Н. Каландадзе, 1987) привел к тому, что эту нишу заполняли самые неожиданные персонажи. Так, с палеоцена по миоцен здесь существовали себекозухии – сухопутные крокодилы с высокой и узкой мордой (предполагают, что их образ жизни напоминал современных комодоских варанов), а в эоцене появились дожившие до плейстоцена фороракосы – гигантские (до 3 м ростом) нелетающие хищные птицы, принадлежащие к журавлеобразным.
В Австралии (Нотогее) формирование травяного биома началось много позже, в неогене; здесь явно сыграл роль дрейф этого континента по направлению от полюса к экватору – в результате значительная часть его территории попала в условия засушливого климата. Основу здешнего сообщества пастбищных млекопитающих составили крупные травоядные сумчатые – кенгуру и вымершие на памяти человека дипротодонты (их иногда, из за двух крупных резцов, не слишком удачно называют «кроликами ростом с носорога»). Как и в древней южноамериканской фауне, здесь отчетливо прослеживается дефицит хищников: известны лишь два крупноразмерных хищных млекопитающих – тилацин (тасманийский сумчатый волк) и древесный тилаколео (которого по аналогии можно назвать «сумчатым леопардом»). Недостаток маммальных хищников возмещался (опять таки как в Южной Америке) за счет рептилий: исполинских варанов мегаланий длиною до 7 м и сухопутных крокодилов, сходных по образу жизни с себекозухиями; хищных нелетающих птиц здесь не возникло, однако некоторые из австралийских страусов, судя по всему, выполняли роль падальщиков.
Третий случай формирования травяного биома – Арктогея. Здесь ситуация осложняется тем, что он формируется на единой таксономической основе (кондиляртровой), но, судя по всему, независимо в Евразии и в Северной Америке. Сообщество пастбищных млекопитающих первоначально составляют непарнокопытные (тапиры, носороги в широком смысле и халикотерии ) и нежвачные парнокопытные (свинообразные и верблюды); чуть позже к ним добавляются примитивные трехпалые лошади и жвачные парнокопытные (олени) (рисунок 55). Помимо потомков кондиляртр нишу крупных растительноядных пытались освоить лишь диноцераты – специализированые потомки каких то крайне примитивных териевых млекопитающих (рисунок 55, б), однако уже в эоцене эта группа полностью вымерла. Единство комплекса «северных» копытных достаточно высоко; самое же интересное состоит в том, что хотя практически все эти группы имеют американское происхождение (они проникали в Евразию через Берингию – области вокруг Берингова пролива, где тогда обсыхали обширные участки шельфа), травяные биомы с их участием в Азии начинают складываться заметно раньше, чем в Америке. В Центральной Азии саванны возникают уже в конце эоцена (появившиеся в это время гигантские безрогие носороги вроде индрикотерия – «гибрид слона и жирафа», самое крупное наземное млекопитающее, 6 м в холке – явно обитали в открытом ландшафте, а не в лесу), тогда как в Америке это происходит в олигоцене. Что касается Африки, то здесь травяных биомов, судя по всему, не существовало до миоцена; парнокопытные и непарнокопытные проникли сюда из Евразии сравнительно поздно, а эндемичные для этого континента хоботные (слоны и мастодонты) были в это время мелкими и чисто листоядными и, видимо, не могли удерживать сукцессию на безлесой стадии.
Что касается хищных млекопитающих, то на Севере они, в отличие от Южных континентов, были только плацентарными: сумчатые вообще существовали здесь очень недолго, и так и не сумели выйти из ниши мелких насекомодных. До того, как здесь появились специализированные плотоядные формы из креодонтов (рисунок 56, а) и современных хищных (Carnivora), в этой роли подвизались своеобразные копытные – мезонихиды (рисунок 56, б в) Мезонихиды были всеядными существами (как полагают, «более плотоядными, чем кабан, но менее плотоядными, чем медведь»); они часто достигали размеров гиены, а эндрюсарх из палеоцена Монголии был крупнейшим наземным хищным млекопитающим – его череп достигает в длину 85 см . Как это ни удивительно, именно от мезонихид ведут свое происхождение китообразные.

РИСУНОК 55. Раннетретичные растительноядные млекопитающие Арктогеи. (а) – кондиляртра (группа, исходная и для «северных», и для «южноамериканских» копытных) Phenacodus; (б) – диноцерат Uiantherium; (в г) – парнокопытные: «жирафовый вердлюд» Aepicamelus (в) и гигантская (высота в плече 2,1 м) свинья Dinochyus ; (д е) – непарнокопытные: халикотерий Moropus (д) и гигантский безрогий носорог индрикотерий (е).

РИСУНОК 56. Раннетретичные хищники Арктогеи. (а) – креодонт Patriofelis длиною около 1,5 м; (б) – собакоподобная мезонихида Synoplotherium; (в) – голова гигантской мезонихиды Andrewsarchus (рядом, в том же масштабе, изображена голова бурого медведя); (г) – диатрима.
До олигоцена ситуация в травяных биомах Арктогеи и Южной Америки развивалась параллельно. И там, и там основными травоядными являлись копытные – потомки различных кондиляртр (на Севере – непарно и парнокопытные, на Юге – «южноамериканские копытные»). И там, и там хищники были явно примитивнее своих жертв (на Юге – сумчатые, на Севере – архаичные всеядные копытные, мезонихиды) – расклад, разительно отличающий палеоген от мезозоя. И там, и там дефицит маммальных хищников возмещался за счет рептилий и птиц: сухопутных крокодилов (на Юге – себекозухии, на Севере – баурузухии) и гигантских нелетающих журавлеобразных (на Юге – фороракосы, на Севере – диатримы). Ситуация эта радикально изменилась, когда на эволюционную сцену вышли современные хищные (отряд Carnivora), и лучше всего это видно как раз по немедленному исчезновению в олигоцене всех этих «эрзац хищников» – всеядных мезонихид, сухопутных крокодилов и диатрим, а также креодонтов (предков карнивор). Интересно, что в то же самое время исчезают и архаичные некопытные растительноядные – диноцераты.
В миоцене единство территорий Северного полушария возрастает: возникает прямой транссредиземноморский контакт между Европой и Африкой, исчезновение Тургайского моря на месте Западно Сибирской низменности облегчает миграции между Европой и Центральной Азией, а главное – возникают открытые ландшафты в чисто лесной доселе Берингии, и эта территория превращается для степных фаун Азии и Америки из «фильтра» в «коридор». С этого времени травяной биом становится фактически единым по всей Арктогее, и при этом каждая из территорий вносит свой вклад в становление его фауны, обретающей уже вполне современные черты.
Из Америки приходят травоядные (в смысле – не листоядные) лошади, из Азии – полорогие (быки и антилопы) , из Африки – хоботные (слоны и мастодонты); вместе с еще некоторыми группами копытных, как «новыми» (жирафы и бегемоты), так и «старыми» (носороги), они формируют так называемую гиппарионовую фауну (гиппарион – одна из трехпалых лошадей). Та же картина и с входящими в гиппарионовую фауну хищными: кошки произошли в Америке, псовые – исходно – тоже, однако стайную социальную организацию (ставшую для этой группы ключевым фактором успеха) приобрели уже в Азии, гиеновые (тогда среди них были не только падалеяды, но и активные хищники типа гепарда) – в Африке. Интересно, что кошки исходно были саблезубыми; позже, в миоцене, возникли кошки современного типа, однако возврат к саблезубости (что, очевидно, дает преимущества при охоте на крупную добычу с особо прочной шкурой) происходил у кошачьих многократно и независимо.
В начале плиоцена (7 8 млн лет назад) самой природой был поставлен грандиозный эволюционный эксперимент: установилась сухопутная связь между Северной и Южной Америками через Панамский перешеек, и их фауны – североамериканская (являющаяся частью Арктогеи) и южноамериканская (пребывавшая весь кайнозой в условиях островной изоляции) – вступили в прямой контакт между собой. Произошло перемешивание фаун: на Севере появились сумчатые, неполнозубые (броненосцы, глиптодонты и наземные ленивцы), кавиморфные грызуны и фороракосы, а на Юге – высшие грызуны (хомякообразные), непарнокопытные (лошади и тапиры), парнокопытные (свинообразные, верблюды и олени), хоботные (мастодонты) и хищные (еноты, куньи, псовые, медведи и кошки). Окончательные результаты Великого американского обмена (как назвал эти события Дж. Симпсон, 1983) оказались, однако, для Севера и Юга весьма различными. Североамериканская фауна просто напросто обогатилась тремя экзотическими «иммигрантами» – опоссумом, девятипоясным броненосцем и древесным дикообразом, – тогда как на Юге произошла настоящая катастрофа, почище любых астероидных импактов: здесь полностью вымер весь пастбищный комплекс из «южноамериканских копытных», гигантских кавиморфных грызунов, хищных сумчатых и фороракосов, не выдержавших конкуренции с высшими копытными и карниворными хищниками (рис 57). Надо полагать, что судьба австралийских сумчатых и однопроходных – возникни у этого континента прямой сухопутный контакт с Азией – была бы столь же незавидной... Вообще в истории Великого американского обмена легко усмотреть прямые (и печальные) аналогии с человеческой историей: вспомним, чем обернулся «контакт» с европейской цивилизацией для древних самобытных культур доколумбовой Америки и Черной Африки.
РИСУНОК 57. «Великий американский обмен». Диаграмма замещения «южных» копытных на территории Южной Америки: в прямоугольниках – число родов, на шкале слева – ярусы южноамериканской региональной шкалы.
Кстати, раз уж речь зашла о человечестве... Согласно современным взглядам, человек (как биологический вид) возник именно в травяном биоме: наши обезьяньи предки входили в состав гиппарионовой фауны. Первые гоминиды появились в конце миоцена (5 6 млн лет назад) на территории Восточной Африки. В прежние годы полагали, что гоминиды обособились среди прочих человекообразных обезьян много раньше, в среднем миоцене (12 15 млн лет назад), и не в Африке, а в Азии (тогда среди прямых предков человека числили индийского рамапитека). Сейчас, однако, полагают, что многие крупные миоценовые приматы (азиатские рамапитек и сивапитек, африканский кениапитек, южноевропейский оранопитек) приобрели «человеческие черты» строения параллеоьно с гоминидами. Палеоантрополог Р. Фоули (1990) пишет: «Рамапитек, после того как ему было отказано в принадлежности к гоминидам, пополнил ряды избранного общества несостоявшихся предков человека, каждый из членов которого обладал, как поначалу полагали, какой нибудь уникальной чертой гоминид»; очевидно, мы в очередной раз имем дело с процессом, который – по аналогии – можно назвать гоминизацией приматов.
Первые гоминиды – австралопитеки – были небольшими прямоходящими существами (весом 25 50 кг); самцы их были почти вдвое крупнее самок – этот резкий половой диморфизм наводит на мысль о том, что они, подобно многим современным приматам, передвигались стаями, не образуя постоянных семей. Около 2,5 3 млн лет назад среди австралопитеков обособились две ветви – робустная (с массивным скелетом, крупными зубами и сильно выступающей челюстью) и грацильная (с легким сложением и относительно большим объемом черепной коробки); робустные виды (Australopithecus robustus, A.boiseni, A.crassidens) были почти чисто растительноядными, тогда как грацильные (A.afarensis, A.africanus) стали широко использовать животную пищу. Именно среди грацильных австралопитеков обособился 2,5 млн лет назад первый представитель рода Homo – Homo habilis, «человек умелый», названный так за способность к изготовлению каменных орудий (первые гальки со следами обработки найдены в слоях возраста 2,5 2,7 млн лет). Он отличался от австралопитеков возросшим объемом черепной коробки и строением таза, обеспечивавшим более совершенную бипедальность и рождение более «головастых» детенышей.
Итак, человек появился в плиоценовой африканской саванне с ее сухим жарким климатом и с изобилием как копытных и хоботных, так и питающихся ими крупных хищников. От обезьян (и от прочих животных) человека отличает кожа, практически лишенная волосяного покрова, но снабженная огромным количеством потовых желез. Уровень потоотделения у человека во много раз превосходит все, что известно в животном мире, и служит чрезвычайно эффективным механизмом теплосброса (за что, правда, приходится платить жесткой связью с источниками воды). По мнению Р. Фоули (1990), именно этот терморегуляторный механизм плюс прямохождение (вертикально стоящий человек получает на треть меньше энергии от солнечных лучей, чем четвероногое животное) позволили первым людям занять в сообществе саванны совершенно уникальную экологическую нишу «полуденного хищника», выйдя из под безнадежной конкуренции с крупными кошками, которые активны в сумерках, а днем спят.
Первые люди были, судя по всему, не столько охотниками, сколько падалеядами, вроде гиен. Впервые появившиеся каменные орудия были наиболее полезны именно для разделки туш очень крупных толстокожих млекопитающих. «Такие звери, обычно неуязвимые для хищников, часто лежат нетронутыми некоторое время после смерти, т.к. другие животные не могут разорвать их кожу и добраться до мяса. Вполне вероятно, что при помощи каменных орудий гоминиды могли первыми начать разделку таких туш и, таким образом, [...] одерживали верх в конкуренции с другими животными, питавшимися падалью» (Р. Фоули, 1990). Каменные орудия, возможно, выполняли ту же функциональную роль, что и огромные клыки саблезубых кошек – их многие палеонтологи считают трупоедами.
Одна из форм внутривидовой кооперации, отличающей человека от прочих животных (включая приматов) – способность делиться пищей; ее считают одной из фундаментальных черт человеческого общества, возникшей (по археологическим данным) уже у плиоценовых гоминид. Видимо, это есть результат разделения труда: вследствие полового диморфизма самцы и самки порознь занимаются поисками пищи (первые – животной, вторые – растительной), после чего следует неким образом поделить собранную добычу. В дальнейшем возникает необходимость в специальном месте, где этот дележ и происходит – иными словами, в жилище. Судя по всему, жилище, разделение труда и дележ пищи возникают уже на самых начальных этапах эволюции человека. Другой комплекс характерных для людей поведенческих реакций связан с заботой о потомстве. Потомство человека зависит от других людей (в первую очередь – от родителей) много дольше, чем у любого другого примата. Одно из следствий этого – высокая степень взаимозависимости человеческих индивидуумов; это касается не только детей, но и самих взрослых, которых объединяет присутствие малышей, нуждающихся в заботе. Доказательством замедленного созревания детенышей мог бы служить характер прорезывания зубов – и ранние гоминиды действительно демонстрируют это свойство. Все это приводит к тому, что основой человеческого поведения становится кооперация между индивидуумами.
Другая (в некотором смысле зеркальная) черта, отличающая гоминид – категорическая неспособность ужиться со своей «дальней родней»: виды рода Homo сугубо аллопатричны, т.е. не сосуществуют сколь нибудь продолжительное время. В раннем плейстоцене (1,6 млн. лет назад) на смену «человеку умелому» пришел «человек прямоходящий» – Homo erectus, которого прежде называли питекантропом; именно в это время полностью вымирают австралопитеки. Примерно 1,2 1,0 млн лет назад «человек прямоходящий» вышел за пределы Африки и заселил южную Азию и Европу, а 0,4 млн лет назад исчез, освободив место для «человека разумного» – Homo sapiens. Термином «архаичные» Homo sapiens обозначают людей, живших в интервале 300 40 тыс лет назад; наиболее известны из них европейские неандертальцы (возраст 70 30 тыс лет) – низкорослые, массивного сложения, с выступающей челюстью, но с мозгом даже более объемистым, чем у современного человека. Homo sapiens современного типа (кроманьонец) появился около 100 тыс лет назад – и опять в Восточной Африке; он заселил Европу 30 40 тыс лет назад и вытеснил неандертальца (злые языки бестактно уточняют – «съел»), практически не смешиваясь с ним.
Вопрос о разумности видов Homo, предшествующих нашему, сводится лишь к спору о терминах («А что есть разум?..»). Во всяком случае, хабилисы уже изготавливали каменные орудия (2,5 млн лет назад), поздние питекантропы пользовались огнем (0,5 млн лет назад), а неандертальцы хоронили своих умерших, совершая сложные погребальные обряды, и создавали произведения изобразительного искусства.
Вот замечательный сюжет для любителей так называемой «альтернативной истории»: 100 тысяч лет назад Африка оказывается начисто изолированной от остального мира (как это действительно произойдет в будущем, через пару миллионов лет – когда до конца раскроется Красноморский рифт), и на планете возникают две цивилизации – кроманьонская в Африке и неандертальская в Евразии. И как знать – может быть, флегматичные неандертальцы создали бы что нибудь более пристойное, нежели то, что мы видим вокруг себя... Впрочем, вряд ли; скорее всего, дело и тут закончилось бы кроманьонской конкистой с «окончательным решением неандертальского вопроса».
История Земли и жизни на ней. Еськов К.Ю., Экспериментальное учебное пособие для старших классов., М.: МИРОС, 1999.
Содержание урока
 конспект урока
опорный каркас
презентация урока
акселеративные методы
интерактивные технологии
Практика
задачи и упражнения
самопроверка
практикумы, тренинги, кейсы, квесты
домашние задания
дискуссионные вопросы
риторические вопросы от учеников
Иллюстрации
аудио-, видеоклипы и мультимедиа
фотографии, картинки
графики, таблицы, схемы
юмор, анекдоты, приколы, комиксы
притчи, поговорки, кроссворды, цитаты
Дополнения
рефераты
статьи
фишки для любознательных
шпаргалки
учебники основные и дополнительные
словарь терминов
прочие
Совершенствование учебников и уроков
исправление ошибок в учебнике
обновление фрагмента в учебнике
элементы новаторства на уроке
замена устаревших знаний новыми
Только для учителей
идеальные уроки
календарный план на год
методические рекомендации
программы
обсуждения
Интегрированные уроки конспект урока
опорный каркас
презентация урока
акселеративные методы
интерактивные технологии
Практика
задачи и упражнения
самопроверка
практикумы, тренинги, кейсы, квесты
домашние задания
дискуссионные вопросы
риторические вопросы от учеников
Иллюстрации
аудио-, видеоклипы и мультимедиа
фотографии, картинки
графики, таблицы, схемы
юмор, анекдоты, приколы, комиксы
притчи, поговорки, кроссворды, цитаты
Дополнения
рефераты
статьи
фишки для любознательных
шпаргалки
учебники основные и дополнительные
словарь терминов
прочие
Совершенствование учебников и уроков
исправление ошибок в учебнике
обновление фрагмента в учебнике
элементы новаторства на уроке
замена устаревших знаний новыми
Только для учителей
идеальные уроки
календарный план на год
методические рекомендации
программы
обсуждения
Интегрированные уроки
Если у вас есть исправления или предложения к данному уроку, напишите нам.
Если вы хотите увидеть другие корректировки и пожелания к урокам, смотрите здесь - Образовательный форум.
|














